
Distinguiamo innanzitutto due casi: nel caso in cui la mutazione avvenga in una cellula somatica, essa sarà presente in tutte le cellule figlie (nate per mitosi) e comporta anche il rischio che quest'ultime diventino tumorali ma non comportano alcun danno alle cellule sessuali; se invece la mutazione avvenisse nei gameti (o nelle cellule che li generano), essa sarà presente in tutte le cellule dell'organismo col rischio incombente di presentarsi anche negli individui di prossima generazione, causando quindi malattie congenite (ereditarie).

cellula tumorale
Ora, distinguiamo alcuni aspetti delle mutazioni genetiche:
- Le mutazioni genetiche hanno origine da diversi fattori, e quindi suddivise in due categorie:
- Mutazioni spontanee: esse sono in genere poco frequenti, oltre che poco incisive. Sono causate da fattori interni all'organismo (errore nella duplicazione, azione diretta di entità ossidanti molto "aggressive"...) e stanno alla base della variabilità genetica (e quindi legate alla selezione naturale).
- Mutazione indotte: queste sono, invece, di frequenza e pericolosità variabili. Sono causate da agenti esterni all'organismo (che quindi li subisce) di varietà e natura innumerevoli che, nel loro insieme, sono definiti mutageni: data la vastità di questa categoria, essi possono agire anche in modi molto differenti per portare alle stesse conseguenze, possono intaccare il materiale genetico direttamente (agenti fisici) o alterando la funzione di alcune molecole dell'organismo (agenti chimici). Inoltre, è proprio tra i mutageni che rientrano anche i portatori delle patologie più gravi (radiazioni UV, raggi X, raggi gamma, composti radioattivi...).

I raggi X
- Le mutazioni genetiche sono suddivise principalmente in tre macro-categorie a seconda del tipo di modifica del corredo genetico (genoma):
- Mutazioni geniche, ossia che alterano un singolo gene. Data la lunghezza relativamente corta di un gene, esse riguardano la modifica di un numero molto piccolo di nucleotidi; le mutazioni spontanee sono quasi essenzialmente geniche e le mutazioni geniche sono spesso spontanee.
- Mutazioni cromosomiche, ossia che alterano i cromosomi. Riguardano il cambiamento della struttura di uno o più cromosomi, dovuta alla loro rottura e successiva riparazione.
- Mutazioni genomiche, ossia che alterano il genoma. Queste ultime sono devastanti, infatti se ne conoscono poche perchè l'organismo solitamente non sopravvive; esse corrispondono ad una modifica nel cariotipo dell'individuo, ossia all'aggiunta o alla perdita nel numero di cromosomi peculiare della specie. Principalmente, le anomalie genomiche sono caratterizzate da trisomia (3 cromosomi) o da monosomia (1 cromosoma): inoltre, esse sono spesso causa di molti tumori.

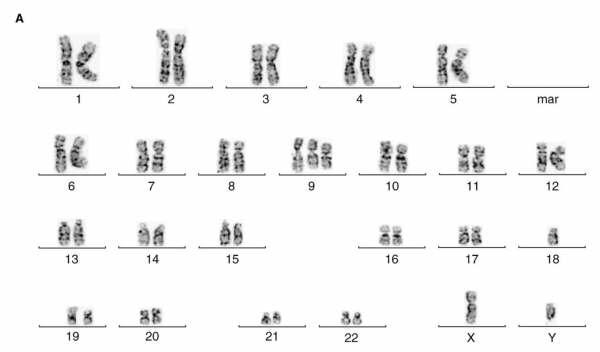
In queste due immagini, si possono notare due mutazioni genomiche diverse riguardanti la stessa coppia cromosomica: nell'immagine più sopra (un cromosoma 18 in più) l'individuo è affetto da trisomia 18, meglio conosciuta come Sindrome di Edwards; nell'immagine meno sopra (un cromosoma 18 in meno) l'individuo è affetto da monosomia 18, ossia la Sindrome di De Grouchy. Sono tutte malattie con conseguenze gravi, se non mortali, per l'organismo.
Sitografia (per maggiori informazioni):
- http://it.wikipedia.org/wiki/Mutazione_genetica











